Family Araliaceae
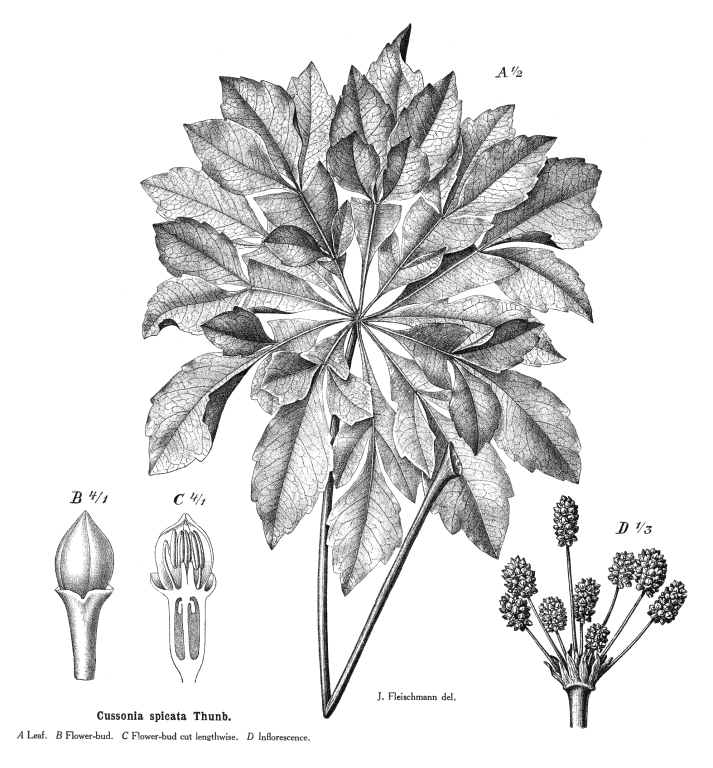
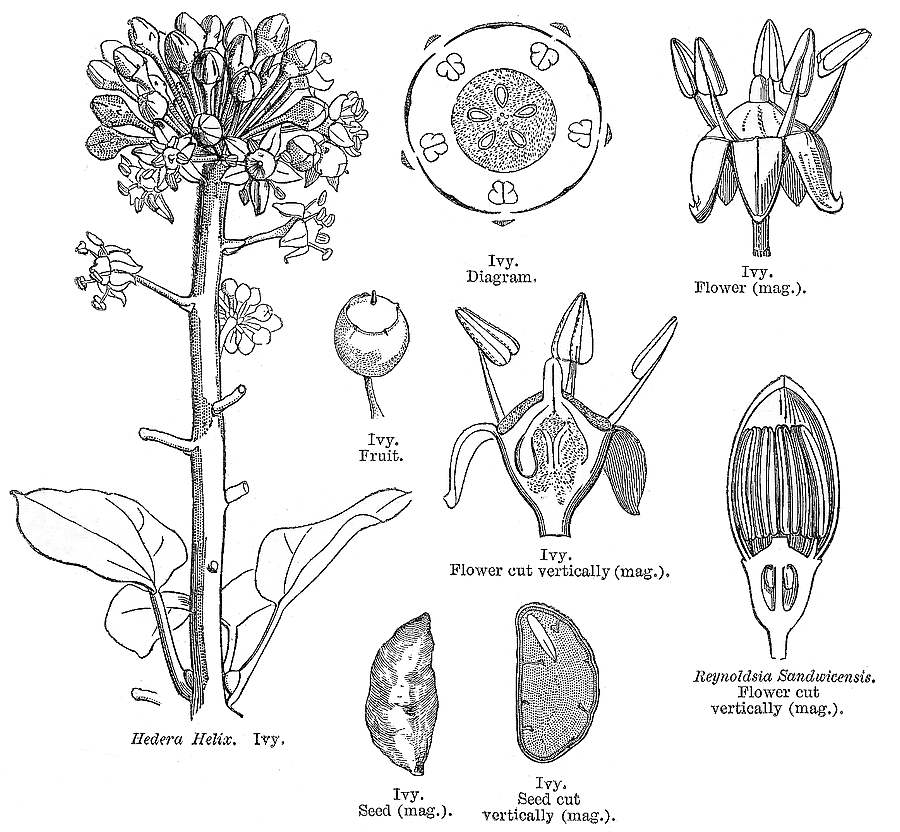
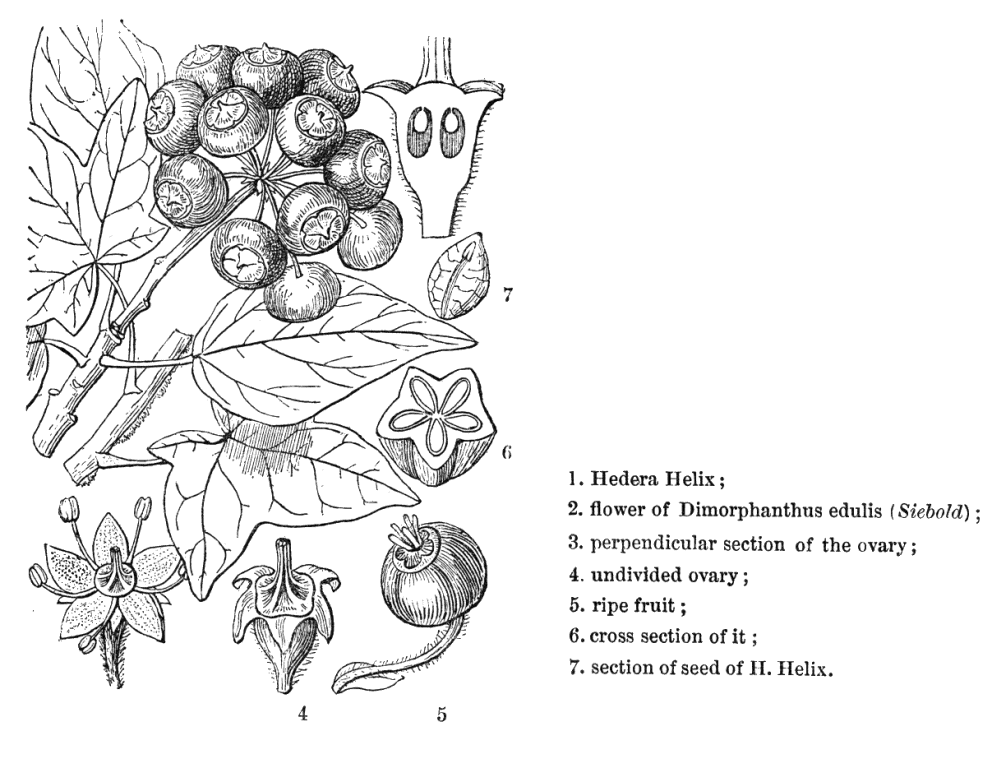
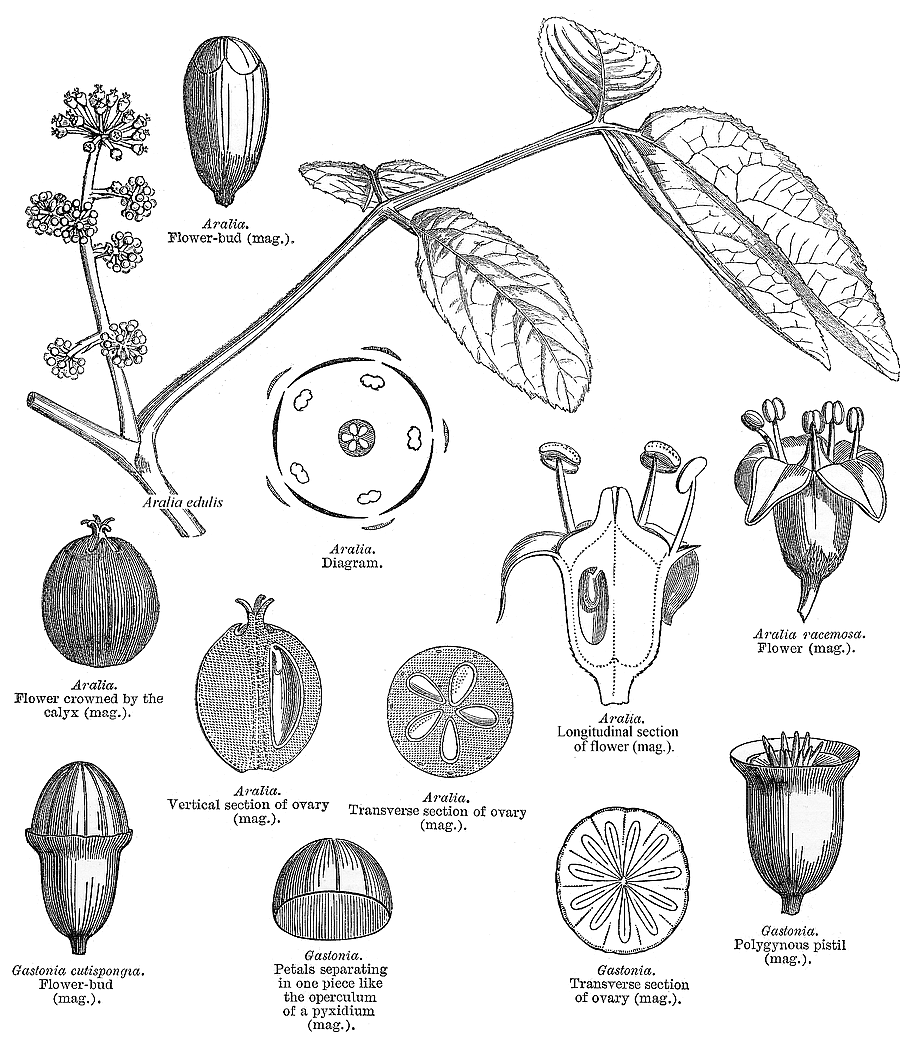
Leaves nearly always alternate (opposite only in Cheirodendron); spiral, or distichous (rarely), or four-ranked (rarely); commonly leathery; petiolate (usually), or subsessile; more or less sheathing (usually), or non-sheathing. Leaf sheaths with free margins. Leaves gland-dotted, or not gland-dotted; aromatic, or without marked odour; simple, or compound; peltate (sometimes), or not peltate; when compound, ternate, or pinnate, or palmate. Lamina when simple, dissected (usually), or entire; pinnatifid, or palmatifid; pinnately veined, or palmately veined. Leaves stipulate, or exstipulate. Stipules when present, intrapetiolar (often adnate to and scarcely distinguishable from the base of the petiole). Vegetative buds scaly. Leaves without a persistent basal meristem; becoming compound from primordial lobes. Leaf anatomy. Stomata present; paracytic (usually), or anomocytic. Hairs present. Complex hairs present (usually), or absent; commonly stellate. Adaxial hypodermis present (commonly), or absent. Lamina with secretory cavities. Secretory cavities containing resin. Minor leaf veins without phloem transfer cells (Aralia, Hedera). Stem anatomy. Secretory cavities present; with resin. Cork cambium present; initially superficial. The cortex without cristarque cells. Nodes multilacunar (often), or penta-lacunar, or tri-lacunar (rarely). Cortical bundles present (commonly), or absent. Medullary bundles present, or absent. Internal phloem absent. Secondary thickening developing from a conventional cambial ring. ‘Included’ phloem absent. Xylem without tracheids; without fibre tracheids; with libriform fibres; with vessels. Vessel end-walls simple (usually), or scalariform (often with few bars), or scalariform and simple. Vessels without vestured pits. Wood parenchyma usually paratracheal (only, often very sparse). Sieve-tube plastids S-type. Reproductive type, pollination. Plants hermaphrodite, or monoecious, or andromonoecious, or gynomonoecious, or dioecious, or polygamomonoecious. Inflorescence, floral, fruit and seed morphology. Flowers aggregated in ‘inflorescences’; in spikes, in heads, and in umbels. Inflorescences terminal, or axillary, or epiphyllous (rarely); umbels or heads, often massed into compound inflorescences; without involucral bracts. Flowers calyptrate (rarely), or not calyptrate; usually more or less 5 merous. Perianth with distinct calyx and corolla, or petaline; (6–)10(–24); 1 whorled, or 2 whorled; isomerous, or anisomerous. Calyx when present, 3–5(–12) (sometimes reduced to small teeth); 1 whorled; when present, polysepalous, or gamosepalous (commonly reduced to a rim); entire, or lobulate, or blunt-lobed, or toothed; often open in bud. Corolla (3–)5(–12); 1 whorled; polypetalous, or gamopetalous (rarely connate at the base or forming a calyptra); calyptrate (rarely), or not calyptrate; imbricate, or valvate; regular. Androecium (3–)5(–12), or 10–100 (the same number as the corolla members, twice the number, or ‘many’). Androecial members free of the perianth; free of one another. Stamens (3–)5(–12), or 10–100; isomerous with the perianth, or diplostemonous, or polystemonous. Anthers dorsifixed; dehiscing via longitudinal slits; introrse; tetrasporangiate. Endothecium developing fibrous thickenings. Microsporogenesis simultaneous. The initial microspore tetrads tetrahedral, or isobilateral. Anther wall initially with one middle layer, or initially with more than one middle layer. Tapetum glandular. Pollen grains aperturate; 3 aperturate (mostly), or 2 aperturate, or 4 aperturate, or 6 aperturate; colporate (mostly), or colpate, or rugate; 3-celled. Gynoecium 2–5(–100) carpelled (i.e., sometimes ‘many’). The pistil 1–200 celled. Gynoecium syncarpous; synovarious to synstylovarious; inferior, or superior (rarely). Ovary 1–200 locular. Locules without ‘false septa’. Epigynous disk present. Styles free, or partially joined. Stigmas wet type, or dry type; papillate; Group II type and Group III type. Placentation apical. Ovules 1(–2) per locule; pendulous; epitropous; with ventral raphe; anatropous; unitegmic; crassinucellate (mostly), or tenuinucellate. Endothelium differentiated. Polar nuclei fusing prior to fertilization. Antipodal cells formed, or not formed (then the three nuclei degenerating early); when formed, 3; not proliferating. Synergids pear-shaped. Hypostase present, or absent. Endosperm formation nuclear. Fruit fleshy, or non-fleshy; indehiscent, or a schizocarp (then cf. Umbelliferae). Mericarps when schizocarpic, 2–5(–100) (?). Fruit when non-schizocarpic, a berry, or a drupe; without fleshy investment. The drupes with separable pyrenes, or with one stone (as many pyrenes as the locules). Gynoecia of adjoining flowers combining to form a multiple fruit, or not forming a multiple fruit. Seeds endospermic. Endosperm ruminate (e.g. Hedera), or not ruminate; oily. Cotyledons 2. Embryo achlorophyllous (2/3). Seedling. Germination phanerocotylar. Physiology, biochemistry. Not cyanogenic. Polyacetylenes recorded (falcarinone). Alkaloids present, or absent. Iridoids not detected (in any of the members screened, discounting a spurious record for Hedera and with Aralidium and Diplopanax excluded). Proanthocyanidins absent. Flavonols present, or absent; quercetin, or kaempferol and quercetin. Ellagic acid absent (5 species, 5 genera). Arbutin absent. Saponins/sapogenins present. Aluminium accumulation not found. Sugars transported as sucrose (Brassaia, Silibertia). Inulin not found (umbelliferose recorded). C3. C3 physiology recorded directly in Hedera. Geography, cytology. Temperate (a few), or sub-tropical to tropical (mainly). Widespread, but especially Indomalaya and tropical America. X = 11, 12(+). Taxonomy. Subclass Dicotyledonae; Tenuinucelli (or borderline, but uncertainties may reflect a few misplaced genera). Dahlgren’s Superorder Araliiflorae; Araliales. Cronquist’s Subclass Rosidae; Apiales. APG (1998) Eudicot; core Eudicot; Asterid; Euasterid II. APG 3 (2009) Order: Apiales. Species 700. Genera 49; Anakasia, Apiopetalum, Aralia, Arthrophyllum, Astrotricha, Boninofatsia, Brassaiopsis, Cephalaralia, Cheirodendron, Cromapanax, Cuphocarpus, Cussonia, Delarbrea, Dendropanax, Eleutherococcus, Fatsia, Gamblea, Gastonia, Harmsiopanax, Hedera, Heteropanax, Hunaniopanax, Kalopanax, Mackinlaya, Macropanax, Megalopanax, Merrilliopanax, Meryta, Motherwellia, Munroiodendron, Myodocarpus, Oplopanax, Oreopanax, Osmoxylon, Panax, Pentapanax, Polyscias, Pseudopanax, Pseudosciadium, Reynoldsia, Schefflera, Sciadodendrom, Seemannaralia, Sinopanax, Stilbocarpa, Tetrapanax, Tetraplasandra, Trevesia, Woodburnia. Diplopanax has been transferred to the cornaceous alliance. This family exemplifies the well known difficulties in distributing certain Dicot families not only between Dahlgren’s Araliiflorae and Corniflorae, but also between the higher level groupings Crassinucelli and Tenuinucelli. This is interesting, given that the latter evidently represent a major divergence in the Dicot line of descent (cf.Young and Watson 1970, Chase et al. 1993). Economic uses, etc. Some cultivated ornamentals, including notable houseplants, e.g. Hedera, Aralia, Polyscias, Schefflera, Fatsia. Ginseng roots from Panax quinquefolius, Chinese rice paper from the pith of Tetrapanax papyriferus. Quotations Sleep thou, and I will wind thee in my arms, . . . . the female ivy so Enrings the barky fingers of the elm (‘Midsummer Night’s Dream’, iv., 1) Oh, falsely they accuse me, |
Support our developers





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}